
Carpet Sea-squirt - Didemnum vexillum
Expand and collapse the sections below by clicking on the title or + / - icons.
Short description of Didemnum vexillum, Carpet Sea-squirt
Pale orange, cream or off-white colonies forming extensive, thin (2-5 mm) sheets; can form long pendulous outgrowths. Firm, leathery texture and veined or marbled appearance. Numerous small pores in surface close when colony disturbed to produce tiny whitish spots; larger water exits occur at intervals.
Impact summary: Didemnum vexillum, Carpet Sea-squirt
Colonies can overgrow other sessile fauna and occupy substantial proportion of available space. On offshore banks (Eastern USA), very extensive coverage of seabed, potentially smothering species living in gravel and affecting fisheries and shellfish industries. Decrease in brittlestars and sea urchins noted in one area (Netherlands).
Habitat summary: Didemnum vexillum, Carpet Sea-squirt
Recorded in GB only from marinas and adjacent shallow artificial structures. In other areas of introduction, also occurs on natural cobble or gravel seabed to 80m depth, in tide pools on shore, in seagrass beds and on bivalve aquaculture installations.
Overview table
| Environment | Marine |
|---|---|
| Species status | Non-Native |
| Native range | Northwestern Pacific |
| Functional type | Filter-feeder |
| Status in England | Non-Native |
| Status in Scotland | Non-Native |
| Status in Wales | Non-Native |
| Location of first record | Holyhead, Plymouth and Dart Estuary |
| Date of first record | 2005 |
Origin
Native range probably NW Pacific and likely to have spread from Japan, although previously overlooked there.
First Record
Recorded in autumn 2008 from a marina in N. Wales and one in Plymouth; photographic evidence of occurrence in 2005 in a marina in the Dart Estuary (Devon).
Pathway and Method
Association with marinas suggests transfer on hulls of leisure craft. Noted in northern France as early as 1998 and probably spread from France to GB. Movements of aquaculture stock has role in some introductions: occurrences on intertidal oyster trestles in Ireland apparently associated with importation of oysters from France. Ballast water is also a likely pathway of introduction.
Species Status
Established populations found in six marinas: N Wales 1, Devon 1, the Solent (Hampshire) 3 and the Clyde 1. One or a few colonies found in each of three more: Devon 1, the Solent (Hampshire and Isle of Wight) 2. (Records for England and Scotland from unpublished Defra-funded surveys, 2009.) Eradication attempted in the N Wales marina 2009-10.
In Europe, first recorded in the Netherlands (1991), and subsequently in northern France (1998), Ireland (2005) and northern Spain (2008).
Also reported in cool-temperate coastal waters in: Japan (first documented occurrence 1926); east coast of the U.S.A., 1982 (anecdotal evidence to the 1970s); west coast of the U.S.A., 1993 (anecdotal evidence to 1991 for British Columbia, Canada); New Zealand, 2001(where described as new to science by Kott 2002).
Dispersal Mechanisms
Natural dispersal ability appears limited: adult sessile, larva brooded prior to release and non-feeding, with brief free-swimming period (minutes to hours). Occasional dispersal by rafting of colonies with detached substrate possible. Pendulous outgrowths on floating structures can ‘drip’ onto seabed below. Colony fragments can reattach and regrow, might be moved some distance beforehand. Long-distance movements likely to have been with commercial oyster shipments, or with relocation of fouled boats or other floating structures.
Reproduction
Adult colony develops from founding individual by asexual budding process. Growth by budding potentially rapid: small colony fragments can grow six- to 11-fold in 15 days. Zooids hermaphroditic. Sexually-produced embryo brooded in basal colonial tunic over two weeks, nourished by yolk included in the egg, then swimming, non-feeding tadpole larva c. 1.6 mm long released. In NE USA, larval production occurs between 14 and 20°C, with some recruitment continuing, as autumn temperatures fall, to 9-11°.
Known Predators/Herbivores
Degenerating colonies were grazed by gastropod Littorina littorea in a Massachusetts (USA) tide-pool. The gastropods Trivia arctica and Lamellaria sp. observed feeding on D. vexillans in the Netherlands. Grazing by chiton Cryptoconchus porosus, sea-urchin Notechinus albocinctus and cushion star Patiriella regularis observed in New Zealand.
Resistant Stages
No resistant or resting stage documented. Colonies may be present year-round, but regress during cold winter weather.
Habitat Occupied in GB
Recorded in GB only from marinas and adjacent shallow artificial structures usually at depths from 30 to 65m at salinities > 26 ppt and temperatures of -2oC to 24oC. In other areas of introduction, also occurs on natural cobble or gravel seabed to 80m depth, in tide pools on shore, in seagrass beds and on bivalve aquaculture installations.
Recorded in a total of nine marinas in GB, in Anglesey (1), Devon (2), the Solent (Hampshire/Isle of Wight) (5) and the Clyde (1).
In Europe, first recorded in the Netherlands (1991), and subsequently in northern France (1998), Ireland (2005) and northern Spain (2008).
Also reported in cool-temperate coastal waters in: Japan (first documented occurrence 1926); east coast of the U.S.A., 1982; west coast of the U.S.A., 1993; New Zealand, 2001.
Environmental Impact
Capable of forming very large colonies, and likely to have considerable effect on pre-existing sessile hard-surface communities through overgrowth interactions etc. Extreme abundance in Oosterschelde, Netherlands (covering >95% of substratum in some areas) accompanied by dramatic decline in brittlestar Ophiothrix fragilis and sea-urchin Psammechinus miliaris. On pebble gravel bottom of Georges Bank off Massachusetts (where locally covered majority of seabed), significant alteration in species composition of benthos compared to uncolonised, including increases in two polychaete species.
Health and Social Impact
None known.
Economic Impact
Possible impacts on the shellfish industry. Larvae of Bay Scallop Argopecten irradians avoided settlement on D. vexillum, suggesting reduction in suitable settlement area where D. vexillum abundant; result of potential significance to fishery for Sea Scallop (Placopecten magellenaicus) on Georges Bank off Massachusetts, where D. vexillum locally abundant. D. vexillum readily overgrows mussels and aquaculture gear. In the Marlborough Sounds mussel farming area of New Zealand, despite initial concern and substantial expenditure on control measures, effects were less severe than initially feared and farmers subsequently opted for a ‘live with it’ policy, rather than seeking funds for ongoing intensive control measures.

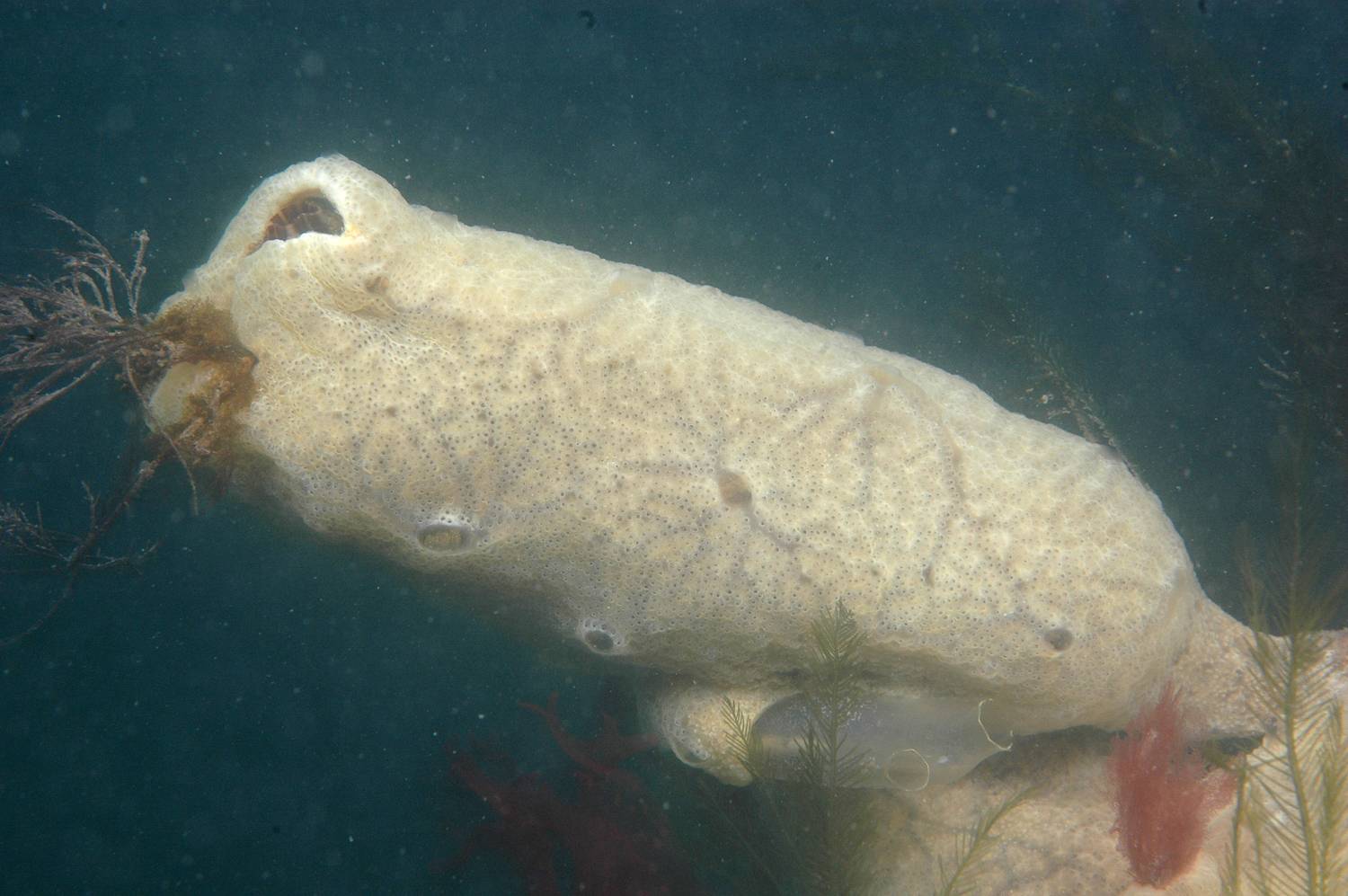

















Identification
Kott, P. (2002) A complex didemnid ascidian from Whangamata, New Zealand. Journal of the Marine Biological Association of the United Kingdom, 82, 625-628.
Lambert, G. (2009) Adventures of a sea squirt sleuth: unraveling the identity of Didemnum vexillum, a global ascidian invader. Aquatic Invasions, 4, 5-28.
Biology, ecology, spread, vectors
Ates, R. (1998) De druipzakpijp, Didemnum lahillei Hartmeyer, 1909 in Zeeland. Het Zeepaard, 58, 101-110.
Breton, G. (2005) Le port du Havre (Manche Orientale, France) et ses peuplements: un exemple de domaine paralique en climat tempéré. Bulletin de la Société Zoologique de France, 130, 381-423.
Carman, M.R. & Grunden, D.W. (2010) First occurrence of the invasive tunicate Didemnum vexillum in eelgrass habitat. Aquatic Invasions, 5, 23-29.
El Nagar, A., Huys, R. & Bishop, J.D.D. (2010) Widespread occurrence of the Southern Hemisphere ascidian Corella eumyota Traustedt, 1882 on the Atlantic coast of Iberia. Aquatic Invasions, 5, 169-173.
Gittenberger, A.(2007) Recent population expansions of non-native ascidians in The Netherlands. Journal of Experimental Marine Biology and Ecology, 342, 122-126.
Griffith K., Mowat S., Holt R.H.F., Ramsay K., Bishop J.D.D., Lambert G.,Jenkins S.R. (2009). First records in Great Britain of the invasive colonial ascidian Didemnum vexillum Kott, 2002. Aquatic Invasions, 4, 581-590.
Minchin, D. & Sides, E. (2006) Appearance of a cryptogenic tunicate, a Didemnum sp. fouling marina pontoons and leisure craft in Ireland. Aquatic Invasions, 1, 143-147.
Valentine, P. C., Carman, M. R., Blackwood, D. S. & Heffron, E. J. (2007) Ecological observations on the colonial ascidian Didemnum sp in a New England tide pool habitat. Journal of Experimental Marine Biology and Ecology, 342, 109-121.
Valentine, P.C., Carman, M.R., Dijkstra, J. & Blackwood, D.S. (2009) Larval recruitment of the invasive colonial ascidian Didemnum vexillum, seasonal water temperatures in New England coastal and offshore waters, and implications for spread of the species. Aquatic Invasions, 4, 153-168.
Management and impact
Denny, C.M. (2008) Developement of a method to reduce the spread of the ascidian Didemnum vexillum with aquaculture transfers. ICES Journal of Marine Science, 65, 805-810.
Lengyel, N.L., Collie, J.S. & Valentine, P.C. (2009) The invasive colonial ascidian Didemnum vexillum on Georges Bank - Ecological effects and genetic identification. Aquatic Invasions, 4.
Morris, J.A.J., Carman, M.R., Hoagland, K.E., Green-Beach, E.R.M. & Karney, R.C. (2009) Impact of the invasive colonial tunicate Didemnum vexillum on the recruitment of the bay scallop (Argopecten irradians irradians) and implications for recruitment of the sea scallop (Placopecten magellanicus) on Georges Bank. Aquatic Invasions, 4, 207-211.
Valentine, P.C., Collie, J.S., Reid, R.N., Asch, R.G., Guida, V.G. & Blackwood, D.S. (2007) The occurrence of the colonial ascidian Didemnum sp on Georges Bank gravel habitat - Ecological observations and potential effects on groundfish and scallop fisheries. Journal of Experimental Marine Biology and Ecology, 342, 179-181.
General
USGS-WHSC: http://woodshole.er.usgs.gov/project-pages/stellwagen/didemnum/htm/page32.htm
Biosecurity Magazine - Issue 86, 15 Sept. 2008 (viewed online 20/5/2010):
http://www.biosecurity.govt.nz/biosec/pubs-news/pubs/biosecurity/issue-86/didemnum
Spotted this species?
Distribution map
View the Distribution map for Carpet Sea-squirt, Didemnum vexillum from NBN Atlas
Risk assessment
Risk assessment for Carpet sea-squirt. See a full list of non-native species Risk assessments.
ID Sheet
ID Sheet for Carpet sea squirt. See a full list of non-native species ID Sheets.
Links to management practices for Didemnum vexillum. See all management information.